
tags: evolution, birds, orioles, Icterus, research

"Oriole."
Image appears here with the kind permission of the photographer, Pamela Wells.
[Larger image].
![]()
I often think about differences in morphological and behavioral traits in closely-related species and wonder whether the speed and character of changes in these traits reveal anything about the evolutionary relationships between taxa. For example, in birds, both visual and auditory cues, such as plumage and song patterns, are essential for identifying members of their own species. However, these phenomena have rarely been systematically examined using closely related species.
To investigate the rate of and types of changes for these traits in closely-related birds, Jordan Price and his colleagues, Nicholas Friedman and Kevin Omland, analyzed plumage color and song patterns in the Icterini, the New World blackbirds, an avian group that includes both the orioles, Icterus, and the oropendolas, Psarocolius and Ocyalus. They then compared their oriole data to a molecular phylogeny for these birds and were surprised to discover that evolutionary plasticity for plumage color and song patterns are similar to each other.
That the rates of behavioral and morphological changes should be different from each other makes intuitive sense; for example, behavioral traits that are influenced by learning, such as birdsong, might be expected to show more plasticity over time than morphological features, such as plumage color patterns, because song varies between individuals of the same species and is known to change rapidly over time due to cultural evolution.
However, previous research has shown that many features of oropendola song are surprisingly conservative and can even be used to identify species with surprising accuracy. In contrast, another study revealed that plumage patterns in orioles is very changeable, with distantly-related species within the group often showing remakable similarities in plumage color and patterns. This interesting conundrum led Price and his colleagues to ask if oriole song evolved in a conservative manner, as previously found for the songs of oropendolas, or if it evolved through repeated convergence and reversals, as seen for the plumage color patterns for orioles?
To answer this question, Price and his colleagues identified 26 distinct vocal characters in oriole songs and analyzed 21 of them in all oriole taxa. Using these data, they reconstructed historical changes in song. The resulting tree revealed that oriole song evolved rapidly, showing repeated reversals and convergences in vocal characters. For example, figure 1 shows two song characters that had high (click; fig. 1a) and average (note diversity; fig. 1b) levels of homoplasy in these birds. Homoplasy describes similar characters that are not derived from a common ancestor.
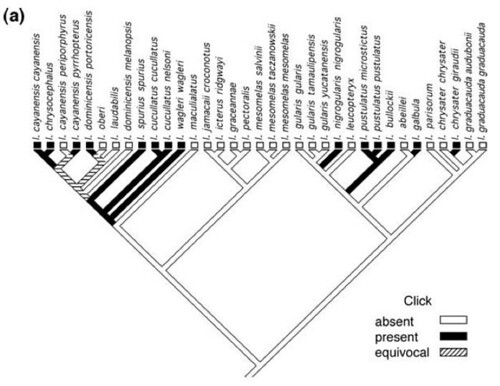

Figure 1: Ancestral state reconstruction of two typical song characters. (a) Click presence or absence has the highest level of homoplasy (CI = 0.11, character 1) and shows at least five independent gains. [Bigger image] (b) Note diversity has close to average homoplasy (CI = 0.22, character 25) and shows multiple examples of both increases and decreases in note diversity, as well as potential reversals [Bigger image]. Character descriptions are in Table 1 [not shown here, available via email]. (MacClade 4.06; ancestral state reconstructions using unordered simple parsimony.)
Price and his colleagues then mapped only those unambiguous changes in oriole song features (figure 2) onto a molecular phylogeny. These data provided strong evidence that song has changed rapidly in the orioles. For example, the Baltimore oriole, Icterus galbula, and black-backed oriole, Icterus abeillei, are genetically very close to each other, yet their songs differed in six of the 21 informative vocal characters that the researchers analyzed, while two subspecies of the streak-backed oriole, Icterus pustulatus pustulatus and I. p. microstictus, which are genetically indistinguishable based on mitochondrial cytochrome-b sequences, produce songs that differ in five vocal characters.
Additionally, you will notice that this tree shows that only 10% of song changes (nine of 83) can be found above the species level, whereas 77% occurred on terminal branches, indicating that song has changed rapidly between closely-related species. You should also note that all of the changes on deeper branches were either convergent with other changes, were reversed on later branches, or both.

Figure 2: Unambiguous evolutionary changes in oriole song reconstructed on the molecular tree. Arrows and character numbers on branches show the gain/increase (up arrows) or loss/decrease (down arrows) in characters [Bigger image]. Note the predominance of changes on terminal and other recent branches of the phylogeny.
Curious to know the relationship between changes in plumage and song patterns with genetics, Price and colleagues compared each of these characters to the molecular distance between species. They found that oriole song patterns appear to be just as homoplastic as oriole plumage patterns (figure 3). This is very different from the evolutionary pattern seen for oropendola song, which revealed that vocal changes accumulated in an almost linear fashion (data not shown) and further, song distance provides a relatively accurate indicator of molecular distance in oropendolas (data not shown).

Figure 3: Plots of (a) oriole song distance and (b) oriole plumage distance as a function of molecular distance (uncorrected p distance based on mitochondrial DNA). Dashed lines illustrating the upper bound nature of both plots were calculated by binning molecular divergence values into 1% increments, selecting the greatest song or plumage distance in each bin (open circles), then calculating a linear regression through these maximal values (see Blackburn et al. (1992); song r =0.74, plumage r =0.80; not evaluated for statistical significance due to nonindependence of data points). The points indicated with an arrow in the lower right of both figures show pairwise comparisons between I. gularis gularis and I. pectoralis, indicating extreme convergence in both overall song pattern and overall plumage pattern. Plumage and molecular divergence values were obtained from Omland and Lanyon (2000) and Omland et al. (1999), respectively.
"These results are surprising because they suggest that there are limits to both song and color variation in orioles (and perhaps in birds in general)," said Price. "Both color patters and song patterns can change pretty rapidly along any particular evolutionary trajectory, but apparently only so far. As a result, within the genus these characters appear to bounce around within a certain, clearly defined, 'character space'."
Price and his colleagues then compared many species of orioles to each other and found that several distantly related species have similar song patterns that arose due to convergent evolution (figure 4). By analyzing the song histories for these birds, Price's team found that none of these birdsongs had similar evolutionary histories (figure 2) and thus, even those that were similar to each other arose independently of each other.
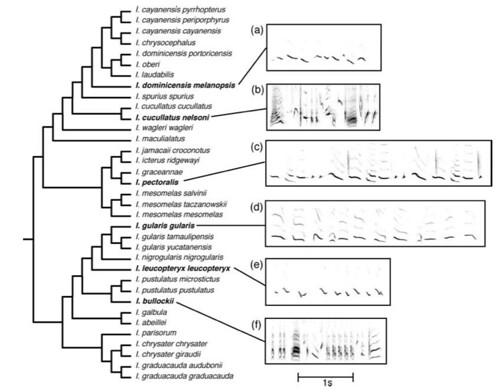
Figure 4: Examples of evolutionary convergence in overall song patterns [Bigger image]. Relationships among taxa are shown in the phylogeny on the left. Songs of I. dominicensis melanopsis, I. cucullatus nelsoni, and I. pectoralis (shown as spectrograms a-c on the right) differ from each other in a variety of features, as do the songs of I. gularis gularis, I. leucopteryx leucopteryx, and I. bullockii (spectrograms d-f). Yet each taxon has a song that is very similar to that of a relatively distantly related taxon (a similar to e; b similar to f; c similar to d). Ancestral state reconstructions indicate that these similarities are explained in part by convergent evolution of multiple, independently derived song features.
"What was especially surprising was that overall song patterns can be really similar between distant relatives," observed Price. "Songs appear to have converged in the orioles much like color patterns have, and not necessarily between the same species. This might be due to the constrained pattern of evolution [described in their paper]. But maybe not. These striking convergences are a little hard to explain."
Source
Song and Plumage in the New World Orioles (Icterus) Show Similar Lability and Convergence in Patterns, by J. Jordan Price, Nicholas R. Friedman, and Kevin E. Omland. Evolution (April 2007) 61-4: 850-863. doi:10.1111/j.1558-5646.2007.00082.x
Jordan Price.
- Log in to post comments
This is cool. I have to read it again and probably another time after that.
As far as the changes in the songs...as I was reading it I was thinking...sounds sort of like language. I wonder if you were a space alien studying humans (hmm and maybe great apes too) and wanted to study the different kinds of humans from different localities, and were to take characters of utterances, focus on particular ones and analyze them (maybe this has been done) what kind of "trees" you might make.
Back to reading!
Chardyspal
the Oriole is one of my favorite birds to watch here in NC, it's a funny little bird